"The Soundtrack of My Life" is a concept that reflects the idea of personal experiences, memories, and emotions being tied to specific songs or music. It often involves compiling a list of songs that resonate with significant moments in someone's life, such as themes of love, loss, joy, and nostalgia. Some people create playlists or write essays that explore how these songs relate to their life stages, relationships, challenges, and triumphs.
"The Temptations in a Mellow Mood" is an album by the iconic Motown band The Temptations, released in 1967. This album showcases a more relaxed and mellow sound compared to their typical energetic performances. It features smooth renditions of classic songs, allowing the group's harmonies and vocal talents to shine through in a softer context. The album includes several covers and original songs that reflect themes of love and yearning, with a blend of R&B and soul influences.
"Undercovers" is an album by the band Night Shift, released in 2021. Night Shift is known for their blend of various musical styles, and "Undercovers" features a collection of cover songs. The album showcases the band's interpretation of tracks from different genres, highlighting their musical versatility and distinctive sound.
"To Wish You a Merry Christmas" is a traditional Christmas carol that expresses warm wishes for happiness and joy during the holiday season. The song is often sung to convey festive cheer and good tidings, typically in gatherings or celebrations around Christmas time. The melody is simple and joyful, making it easy for groups to sing together. The lyrics often emphasize themes of goodwill, merriment, and the spirit of Christmas.
"Together" is a collaborative studio album by two iconic Motown groups, The Supremes and The Temptations. Released in 1969, the album features a mix of soul and pop songs, showcasing the vocal talents of both groups. The project is notable for its unique blend of the smooth harmonies of the Supremes with the energetic vocal style of the Temptations.
"Triplicate" is a studio album by Bob Dylan, released on March 31, 2017. It is a triple album that features 30 tracks, all of which are covers of classic American songs, predominantly from the early to mid-20th century. The album continues Dylan's exploration of the Great American Songbook, following his previous albums, "Shadows in the Night" (2015) and "Fallen Angels" (2016), which also focused on classic standards.
Truth & Soul is an independent record label and publishing company based in Los Angeles, California, known for its focus on soul, funk, and R&B music. Founded in 2004 by Leon Michels, a musician and producer, Truth & Soul has been instrumental in bringing a contemporary sound to these genres while drawing heavily on classic influences. The label has produced and released music for various artists and projects, including notable acts like El Michel's Affair, Lee Fields, and The Expressions.
"Twelve" is a cover album by American musician Patti Smith, released in 2007. The album features Smith's interpretations of twelve songs by various artists that have influenced her throughout her career. The tracks include reimagined versions of songs by notable musicians such as Bob Dylan, Neil Young, and The Doors. Each song showcases Smith's unique style and vocal delivery, blending her rock roots with poetic lyrics and emotional depth.
The "Crysta-Rhythm" album is a tribute album to the Japanese pop duo Two-Mix, known for their contributions to the J-pop and anime music scenes in the 1990s and early 2000s. Released in 2016, "Crysta-Rhythm" features various artists interpreting and reimagining popular tracks from Two-Mix's catalog. The album showcases the influence that Two-Mix has had on other musicians and highlights their iconic sound through different styles and arrangements.
"Uncovered" is an album by British musician Steve Harley, released in 2014. This album features a collection of cover songs, showcasing Harley's interpretation of various tracks from different artists. The album includes selections that highlight his distinctive style, blending rock and pop elements with his unique vocal delivery. Known for his previous work with the band Cockney Rebel and his solo career, "Uncovered" reflects his diverse musical influences and offers a fresh take on well-known songs.
"Wallflower" is an album by Canadian jazz pianist and vocalist Diana Krall. Released in 2015, it features a collection of covers of classic pop and rock songs, reimagined in Krall's signature jazz style. The album includes interpretations of tracks originally performed by artists such as The Eagles, Bob Dylan, and Harry Nilsson, among others.
The term "D-ring" can refer to different things depending on the context. Here are a few common interpretations: 1. **Hardware**: In general hardware terminology, a D-ring is a metal or plastic fastening device shaped like the letter "D." It's often used in applications such as securing straps, tents, cargo, and climbing gear, as it provides a sturdy point for attaching ropes or straps.
"Utaite Myōri: Sono Ichi" is a Japanese anime series that explores themes related to music and performance. While it may not be widely known or as mainstream as other titles, it typically features characters who are involved in singing or music-related competitions, often highlighting their struggles, aspirations, and the impact of music on their lives. The term "Utaite" generally refers to singers, particularly those who cover songs, often from Vocaloid or popular anime.
"Vanilla Ice Is Back!" is a reality television show that premiered in 2016 featuring the rapper Vanilla Ice, who gained fame in the early 1990s with his hit song "Ice Ice Baby." The show follows Vanilla Ice as he endeavors to revitalize his music career while also engaging in various projects, including home renovation. The series highlights his personal life, friendships, and efforts to navigate the modern music industry.
"Utahime" can refer to a couple of different things depending on the context. 1. **Utahime in Music**: In the realm of Japanese pop culture, "Utahime" can refer to a "singer" or "vocalist" as the term itself translates to "song princess" or "singing woman" in Japanese. It is often used to describe female singers, especially those who have a significant impact or popularity.
"Versão Acústica 3" is the title of an acoustic album by the Brazilian rock band **Fresno**. Released in 2017, it features acoustic renditions of some of the band's popular songs, showcasing their music in a stripped-down format that emphasizes vocals and acoustic instrumentation. The album continues Fresno's exploration of their sound and has been well-received by fans, highlighting the band’s emotional lyrics and melodies.
"Vintage Dead" is a novel written by author Robert E. Howard, originally published in 1930. The book falls within the horror and supernatural fiction genres and showcases Howard's signature style of weaving together adventure and the macabre. The story involves themes that are typical of Howard's work, including elements of mysticism and the supernatural. It typically features a protagonist who encounters ghosts or otherworldly beings and must confront the darker aspects of humanity.
Vocalo Zanmai is a Japanese music project known for its use of Vocaloid technology. The term "Vocaloid" refers to a singing synthesizer application that enables users to create music using synthetic voices, often represented by animated characters. Vocalo Zanmai features various virtual singers and emphasizes collaborative music production, typically involving multiple producers and artists. The project has gained popularity for its catchy songs and engaging visuals, often combining elements of pop, electronic, and sometimes traditional Japanese music.
As of my last update in October 2023, there isn't a widely recognized product, service, or concept specifically called "Voice Nova." It's possible that it could refer to a number of things depending on the context, such as: 1. **Voice Technology**: It might refer to a voice analysis or speech recognition technology, a voice assistant, or a platform related to voice communication.
Pinned article: Introduction to the OurBigBook Project
Welcome to the OurBigBook Project! Our goal is to create the perfect publishing platform for STEM subjects, and get university-level students to write the best free STEM tutorials ever.
Everyone is welcome to create an account and play with the site: ourbigbook.com/go/register. We belive that students themselves can write amazing tutorials, but teachers are welcome too. You can write about anything you want, it doesn't have to be STEM or even educational. Silly test content is very welcome and you won't be penalized in any way. Just keep it legal!
Intro to OurBigBook
. Source. We have two killer features:
- topics: topics group articles by different users with the same title, e.g. here is the topic for the "Fundamental Theorem of Calculus" ourbigbook.com/go/topic/fundamental-theorem-of-calculusArticles of different users are sorted by upvote within each article page. This feature is a bit like:
- a Wikipedia where each user can have their own version of each article
- a Q&A website like Stack Overflow, where multiple people can give their views on a given topic, and the best ones are sorted by upvote. Except you don't need to wait for someone to ask first, and any topic goes, no matter how narrow or broad
This feature makes it possible for readers to find better explanations of any topic created by other writers. And it allows writers to create an explanation in a place that readers might actually find it.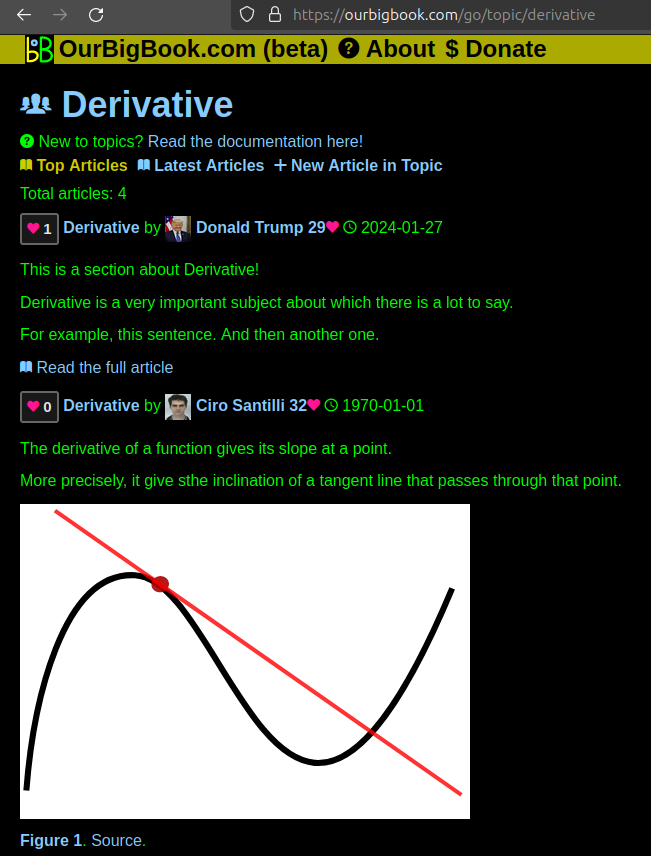
Figure 1. Screenshot of the "Derivative" topic page. View it live at: ourbigbook.com/go/topic/derivativeVideo 2. OurBigBook Web topics demo. Source. - local editing: you can store all your personal knowledge base content locally in a plaintext markup format that can be edited locally and published either:This way you can be sure that even if OurBigBook.com were to go down one day (which we have no plans to do as it is quite cheap to host!), your content will still be perfectly readable as a static site.
- to OurBigBook.com to get awesome multi-user features like topics and likes
- as HTML files to a static website, which you can host yourself for free on many external providers like GitHub Pages, and remain in full control
Figure 3. Visual Studio Code extension installation.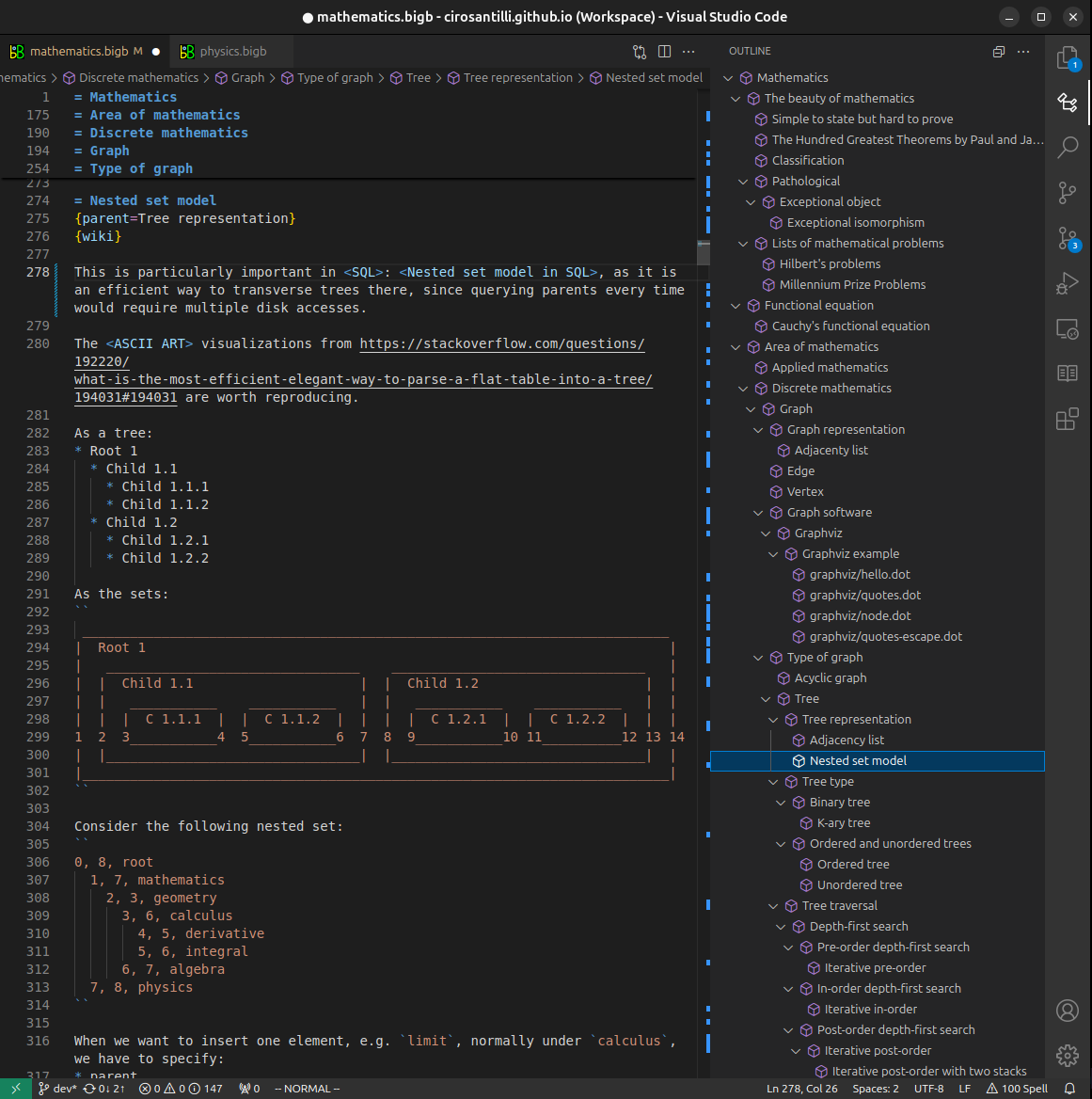
Figure 4. Visual Studio Code extension tree navigation.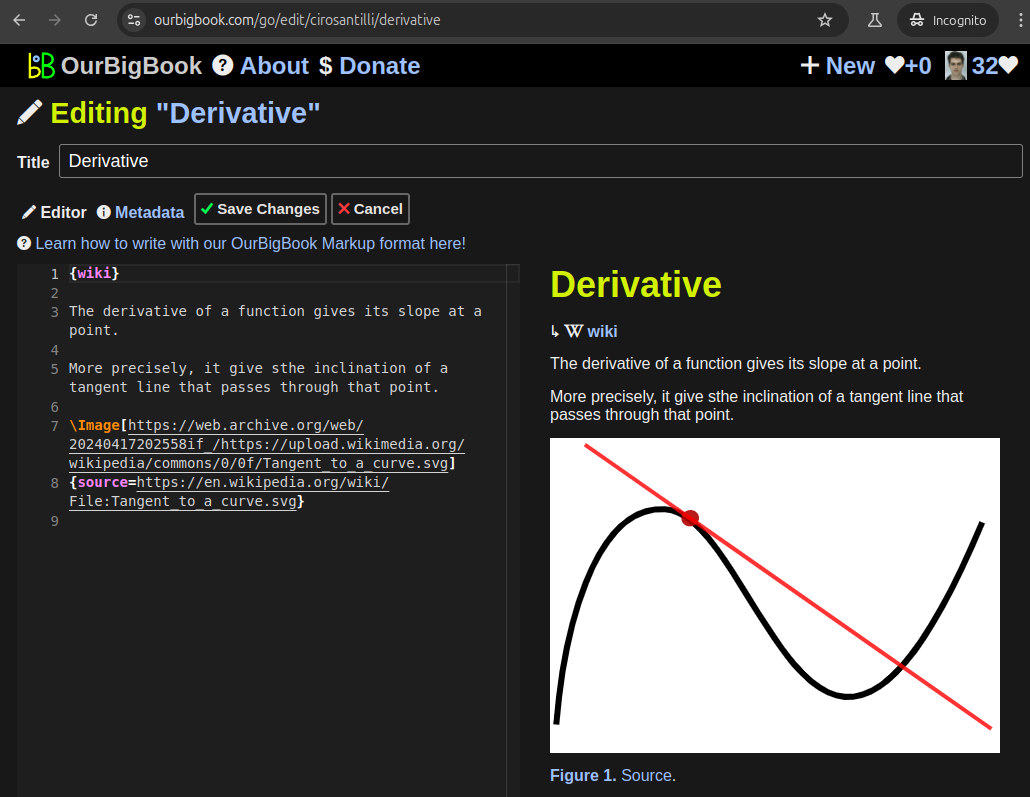
Figure 5. Web editor. You can also edit articles on the Web editor without installing anything locally.Video 3. Edit locally and publish demo. Source. This shows editing OurBigBook Markup and publishing it using the Visual Studio Code extension.Video 4. OurBigBook Visual Studio Code extension editing and navigation demo. Source. 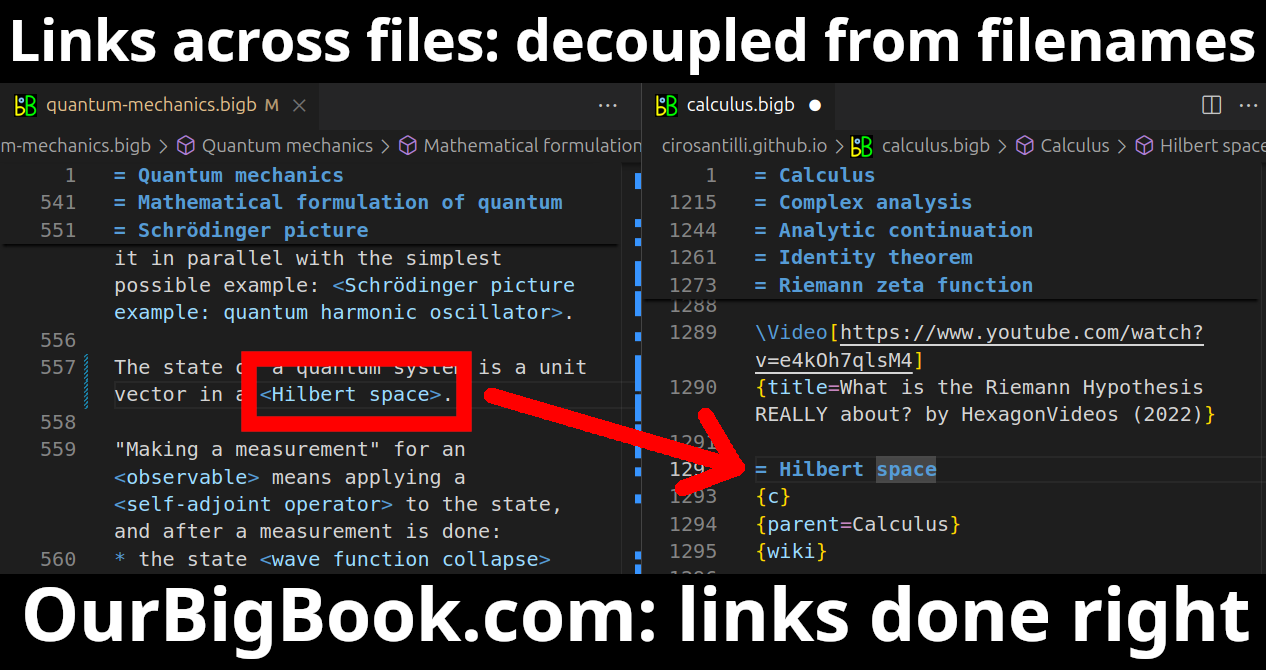
- Infinitely deep tables of contents:


All our software is open source and hosted at: github.com/ourbigbook/ourbigbook
Further documentation can be found at: docs.ourbigbook.com
Feel free to reach our to us for any help or suggestions: docs.ourbigbook.com/#contact